
近日,我校梁祥修教授、四川农业大学马苗苗副教授、崖州湾国家实验室周俭民研究员共同在Science Bulletin上撰写题为“Phosphorylation-dependent Regulation of Plant Heterotrimeric G Proteins: From Activation to Downstream Signaling”的观点文章,综述了磷酸化修饰在G蛋白从激活到下游信号转导过程中的调控功能和机制。基于近年来植物G蛋白研究领域的进展,总结了植物G蛋白独特的激活和信号转导模式。

异源三聚体G蛋白由α、β和γ三个亚基组成,是真核细胞重要的分子开关蛋白。动物G蛋白与细胞表面7次跨膜的G蛋白偶联受体(GPCR)结合,GPCR感知胞外信号后,其胞内结构域发生构象改变,使与Gα结合的GDP分子被GTP替换,G蛋白激活并发生三聚体解离,激活后的Gα与Gβγ通过作用于下游靶标进行信号转导。植物异源三聚体G蛋白在生长发育、氮肥吸收利用、作物产量、非生物胁迫响应以及病原物入侵等过程中都发挥关键作用。但长期以来,对植物G蛋白的激活及下游信号转导分子机制的认识相对欠缺。
单次跨膜的受体类激酶(RLK)和受体类蛋白(RLP)是植物细胞表面最主要的受体类型,其主要依赖磷酸化修饰进行信号转导。近年来研究发现,植物G蛋白与RLK或RLP受体偶联,以此感知胞外信号,并通过作用于下游多个家族的蛋白激酶进行信号转导。因此磷酸化修饰在植物G蛋白的激活和下游信号转导过程中都发挥关键作用。
植物异源三聚体G蛋白的激活存在多个层次的调控。具有GTP水解酶加速活性的RGS1蛋白存在于多数双子叶植物中,RGS1与Gα互作并抑制G蛋白处于静息状态。RLK/RLP受体(如,植物识别细菌鞭毛蛋白的免疫受体FLS2)感知胞外信号后,通过磷酸化修饰RGS1,调控RGS1蛋白的内吞,从而调节RGS1与Gα的互作关系。Gα自身也受到磷酸化修饰,该磷酸化也能够影响RGS1-Gα互作。RLK/RLP受体通过调控RGS1与Gα的互作,从而调节植物G蛋白的激活(图1)。此外磷酸化修饰还通过影响RGS1的酶活性和G蛋白亚细胞定位等方式影响G蛋白的功能。
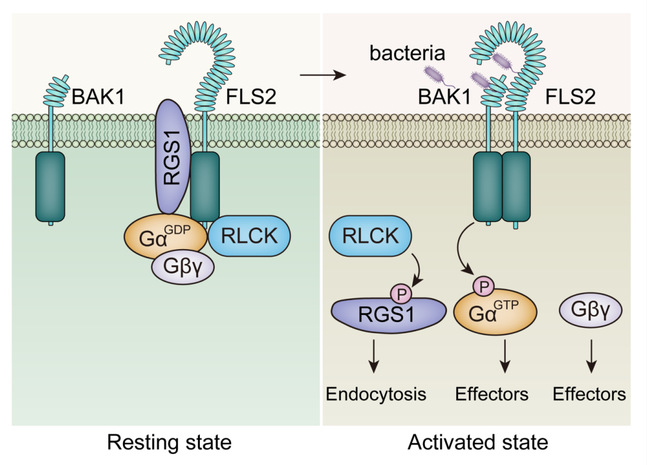
图1. RLK/RLP受体通过调控RGS1和Gα蛋白的磷酸化状态调节植物G蛋白激活
在G蛋白下游信号转导方面,G蛋白与多个蛋白激酶家族存在相互作用,包括细胞质类受体激酶(RLCK)、MLK激酶、MAPK激酶等。这些激酶家族在许多生物学过程中都发挥重要功能,G蛋白通过调控这些激酶的稳定性或活性,影响其介导的下游信号途径,从而实现G蛋白下游信号转导(图2)。
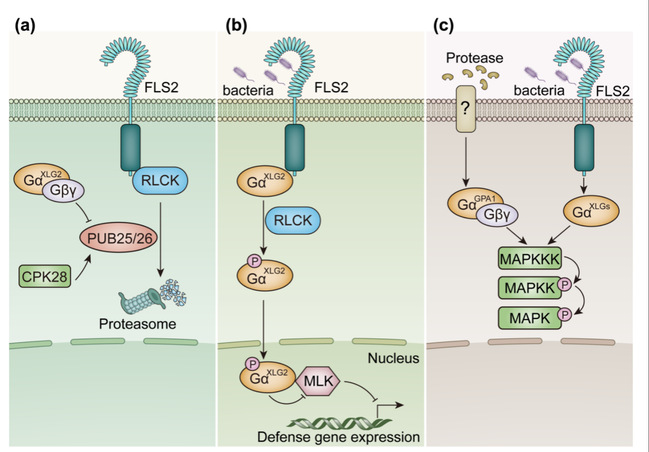
图2.植物G蛋白通过调控多个蛋白激酶家族传递下游信号
本文最后对植物G蛋白研究方向尚未解决的关键科学问题和未来研究方向进行了展望。对单子叶植物G蛋白的激活机制的探索,以及鉴定G蛋白新的下游靶标,将有助于系统认识植物G蛋白的激活机制及其下游信号转导网络,从而更好地理解植物如何通过精细的信号调控网络来适应环境变化。为基于植物异源三聚体G蛋白,培育高产抗逆的作物品种提供理论基础。
华南农业大学梁祥修教授、四川农业大学马苗苗副教授和崖州湾国家实验室周俭民研究员是本文共同通讯作者。该文章获得国家自然科学基金(32300260, 32270282)和双一流学科建设项目(2021B1056400)的支持。
相关论文信息:https://www.sciencedirect.com/science/article/pii/S2095927324003165?via%3Dihub=
文图/生命科学学院
